In all domains of life, transcription elongation factors bind to the transcribing RNA polymerase to control gene expression. Many of these factors are recruited in a sequence-specific manner, but catching a moving target is no easy feat. Most commonly, elongation factors identify the target sequence within the newly synthesized RNA and then load onto RNA polymerase – this mode of recruitment, exemplified by CPSF in eukaryotes and the rRNA antitermination complex in prokaryotes, affords a relatively broad time window available for binding of a factor to RNA polymerase.
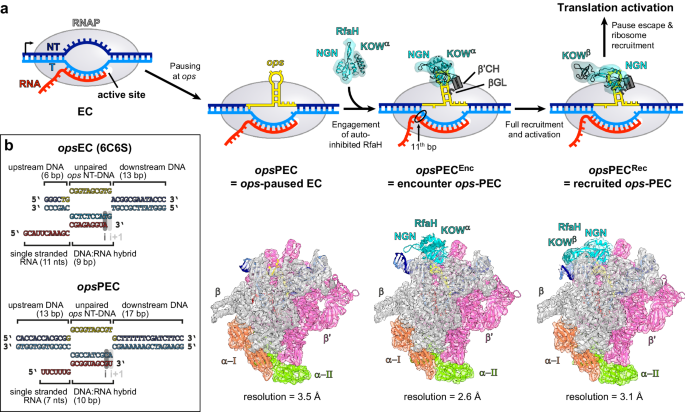
In contrast, the bacterial transcription antiterminator RfaH uniquely recognizes its short target sequence, called ops, in the non-template DNA strand exposed on the surface of the transcription elongation complex. The RfaH target would be available for RfaH binding only for a fraction of a second unless RNA polymerase pauses to extend the time window for RfaH recruitment, and ops has been shown to induce pausing. Timely and efficient RfaH recruitment is critical for bacterial fitness and pathogenesis, as well as for the horizontal spread of antibiotic resistance - RfaH activates expression of long operons that encode adhesins, capsules, lipopolysaccharide core, toxins, and conjugal transfer systems by recruiting the pioneering ribosome and preventing premature termination of RNA synthesis. But each of these operons has only a single ops element in an untranslated leader region, making RfaH loading once-in-a-lifetime opportunity for an mRNA to be synthesized in its entirety.
At the same time, RfaH recruitment must be tightly controlled to prevent mis-regulation of housekeeping genes transcribed by RNA polymerase bound to NusG, an essential paralog of RfaH, because RfaH binds to RNA polymerase much tighter than NusG. To avoid off-target recruitment, free RfaH is autoinhibited by its C-terminal KOW domain, which folds as an α-helical hairpin and masks the RNA polymerase-binding site located on the N-terminal NGN domain. Interactions with the ops DNA trigger RfaH activation, which is accompanied by the release of KOW to enable NGN binding to RNA polymerase; at the same time, KOW completely re-folds into a β-barrel and binds the ribosome to couple transcription to translation. Thus, the ops element must (i) halt RNA polymerase to enable RfaH binding; (ii) trigger RfaH activation, and (iii) delay the RNA polymerase/RfaH complex until the ribosome is recruited.
Here, Zuber et al. obtained a series of cryoEM structures that capture key events along this regulatory pathway. First, they present a structure of the ops-paused complex stabilized in a catalytically inactive pre-translocated state with an 8-nt-long nontemplate DNA hairpin formed on the surface of RNA polymerase. The ops DNA hairpin triggers swiveling of RNA polymerase structural modules to prolong the pause while also presenting multiple recognition determinants to facilitate subsequent binding of RfaH.
Next, they present a structure of an encounter complex in which RfaH, artificially locked in the autoinhibited state by an interdomain crosslink, is bound to the ops-paused RNA polymerase. This structure shows that, prior to domain dissociation, RfaH binds to, and repositions, the ops DNA hairpin to initiate expansion of the transcription bubble and the RNA:DNA hybrid, further stabilizing the paused state. Using the encounter complex as a starting structure, the authors used molecular dynamics simulations to reveal the mechanism by which the DNA hairpin "picks the lock" in RfaH, triggering its activation, whereas RNA polymerase plays a largely scaffolding role.
A structure of activated RfaH fully engaged with RNA polymerase shows how RfaH recruitment stabilizes an even deeper paused state, likely making escape dependent on accessory factors. Indeed, the authors also present a structure that captures an unsuccessful escape attempt of RfaH-bound RNA polymerase which, after extending the RNA by two nucleotides, backtracks to the original position. Biochemical data show that anti-backtracking factors GreA and Mfd facilitate escape and suggest that the pioneering ribosome may play this role in the cell.
Remarkably, this elaborate functional cycle may be widespread in nature. NusG proteins are the only universally conserved transcription factors and many genomes encode specialized NusG paralogs, with up to 20 % of them being potentially metamorphic. Studies of RfaH presented in this work offer a blueprint for deciphering the recruitment and folding switch mechanisms of these ancient and ubiquitous regulators.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in