Eating seasonally: How studying dietary changes can help conservation
Published in Ecology & Evolution

A common adage in conservation is that ‘we cannot conserve what we don’t understand’. It can be surprising then that our knowledge of even iconic species can be incomplete or inaccurate. In the scramble to protected species slipping towards extinction, it is completely understandable that money and effort is directed towards tackling threats head on, but without accurate knowledge conservation programmes can be ineffective. Rigorous research and new technologies can help to reassess received wisdom in ecology and improve conservation.
Species stereotypes
We use the term ‘species stereotype’ to refer to misunderstandings of species’ ecology, such as the habitats that are optimal for breeding and survival, caused by unrecognised research biases [1]. Research can be biased by a whole range of things, including how easy it is to access study areas or observe certain behaviours. As an example, cheetah are widely considered to be savanna specialists, but there is evidence to suggest that they actually do well if not better in thickets [2]. This may be partly because behaviour in closed habitats is harder to observe, and partly because researchers mainly go to savannas to study them in a self-fulfilling prophecy.
It is not easy to bust these stereotypes in any situation, but when a species has undergone range declines it is particularly difficult as we cannot study something where it no longer exists. One way around this is to research the mechanisms behind the impact of the environment on the survival and breeding success of individuals, and how that scales up to population level impacts of decline, recovery and extinction [3]. This mechanistic understanding can be used as evidence to improve conservation programmes and identify suitable habitat even outside of current ranges.
Critically endangered herbivores
In our newly published paper ‘Linking diet switching to reproductive performance across populations of two critically endangered mammalian herbivores’ [4] we apply these principles to eastern black rhino and Grevy’s zebra in Kenya. Specifically, we use the change in what these species eat seasonally to uncover dietary preferences, the impact of seasonal diet changes on their gut microbiome, and ultimately the effect on breeding success. Both species are restricted to tiny pockets of habitat, but studying populations that exist across a rainfall gradient can give insights into their performance in habitats that exist across their historical ranges.
Black rhino are considered to be browsers, and estimations of how many can be supported by a particular habitat are largely based on the density of acacia trees, in the Fabaceae family (legumes), those often flat-topped and thorny trees you see on nature documentaries.

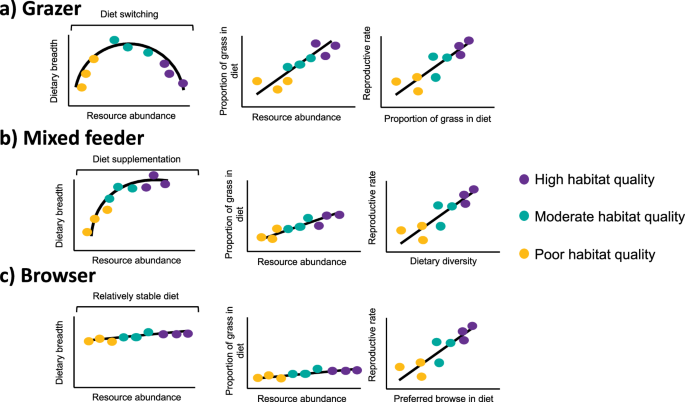
By contrast, Grevy’s zebra are grazers with a preference for grasses (plants in the Poaceae family). We had different predictions for how different guilds of herbivores should vary their diets seasonally, and how this will affect breeding rate (Figure 1).

Figure 1. Predictions for hypothetical large mammalian herbivores with different feeding strategies
We used metabarcoding to study the diets and microbiome of the two species, across three reserves for each. Metabarcoding uses DNA sequencing of a particular gene or region to match the taxa that are present in a particular sample to a library of known sequences. In our case that meant picking up a lot of dung and extracting plant and bacterial DNA. We looked at how diets and microbiome changed with a measure of vegetation greenness, obtained using satellite remote sensing, called NDVI. After rainfall in savanna, vegetation greens up and NDVI increases. In this blog we only discuss the diet results of our research.
What we found
Grevy’s zebra
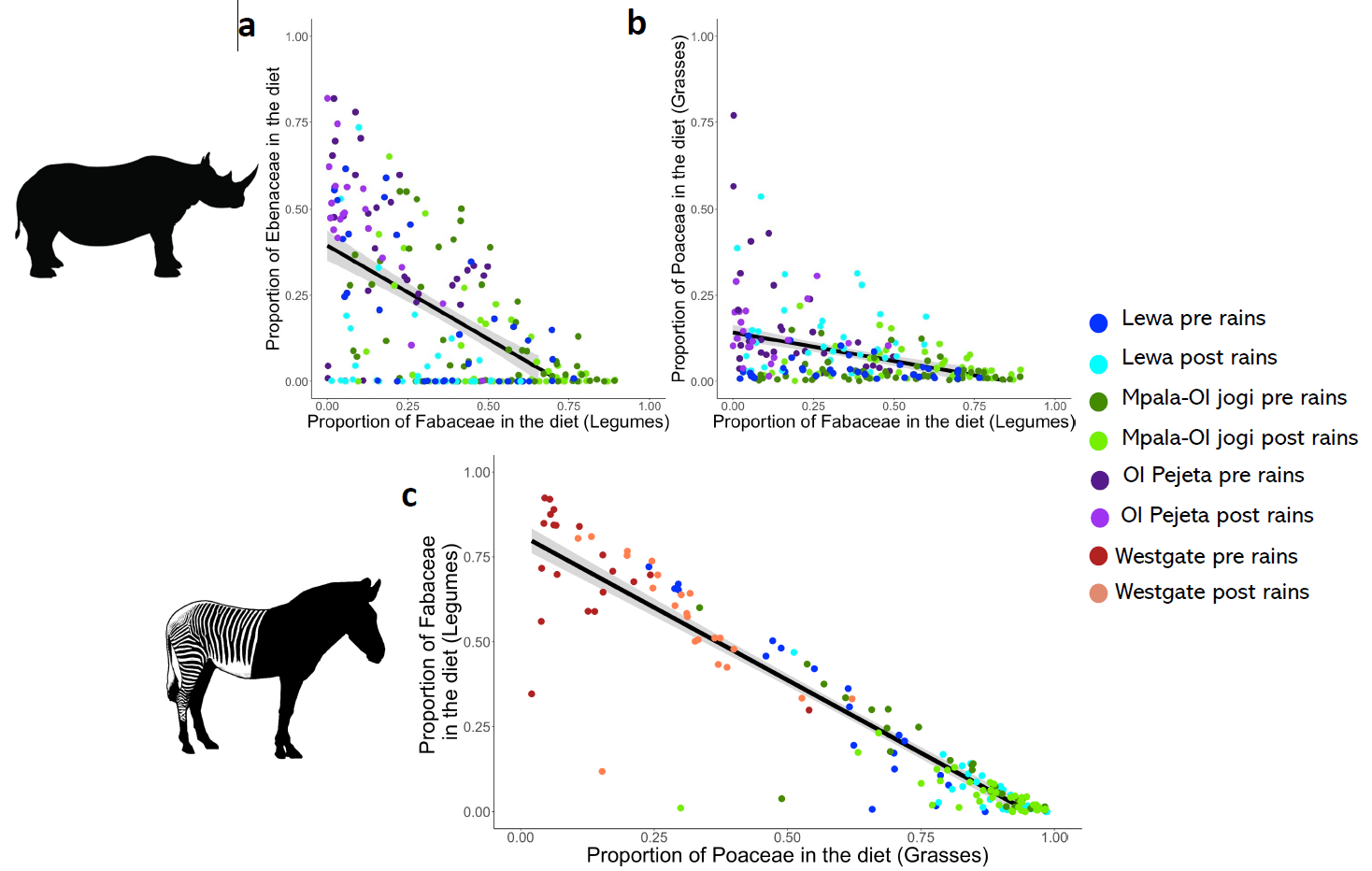
Grevy’s zebra largely conformed to expected patterns, grass is clearly their preferred food. In drier seasons and places, they switched to fallback foods in the Fabaceae family (Figure 2), mostly a genus called Indigofera. The strength of the correlation between the two plant families (Figure 3) suggests that they didn’t consume much else, and that Fabaceae is a much lower value food than grass. Zebra on different reserves have drastically different diets and the very arid Westgate must be a marginal habitat.

Black rhino
The story for black rhino is not so straightforward, but a key result is that they seem to be a less strict browser, and closer to being a mixed feeder, than previously thought. As NDVI increased, rhino consumed less acacia (Fabaceae family) and more grass. This suggests that acacia acts as a fallback food rather than the preferred food that it is widely thought to be. Switching between acacia and grass was not as clear cut as it was for the zebra, partly because all rhino ate some acacia year round, and partly because they also consume plants from other families, including the Ebenaceae family that are particularly common on Ol Pejeta.
Why does all this matter?
In Kenya, neither species can move around freely to find the best habitat. Because of this, we need conscious planning of where these animals should be conserved, and diet is a vital part of this. As the black rhino population continues to expand, current reserves are reaching capacity and conservation managers are continually trying to identify new areas to found populations.
The final link in the chain of our research was tying diet characteristics to breeding. For zebra, we looked at the correlation between the proportion of foals in the population (a high proportion indicates better population performance) and the consumption of Fabaceae, which we have found to be fallback foods (Figure 4). This relationship was not significant, but the analysis was constrained to a coarse scale and small sample size. Our other results do however provide strong evidence that grass availability should be a central consideration for Grevy’s zebra conservation.
For black rhino, females which consume more acacia have larger seasonal diet shifts, and in turn breed more slowly (Figure 4). Dietary switching may therefore indicate marginal habitats where individuals fallback to consume acacia in times of scarcity and can only eat high-value plants at certain times of the year.

There is strong evidence that this is impacting the population-level dynamics that conservation really cares about. Our previous work demonstrated that female black rhino on Ol Jogi exhibit lower breeding and higher mortality rates than Lewa and Ol Pejeta, and the population there is more susceptible to extinction [5]. Acacia makes up a higher proportion of rhino diets on Ol Jogi, and there is much less grass available there year-round. This is not to say that acacia is not important for black rhino, they consume it all year and it is an important buffer against drought, but other plants should be included in habitat suitability assessments.
This work demonstrates how important it is not to rely on received wisdom about what species should be eating and what their favoured habitats are, especially when they have undergone severe range declines. Stereotypes can be busted by new technologies which make it possible to study the diets and physiology of endangered species more accurately. As the global environment changes rapidly, we need this to make well-informed decisions about where and how to conserve even our most iconic species.
References
[1] Britnell, J.A., Lewis, R.N., Elsner-Gearing, F., Harvey, N., Stanbrook, E. and Shultz, S., 2021. Species stereotypes as a result of unconscious research biases compromise conservation efficacy. Biological Conservation, 261, p.109275.
[2] Bissett, C. and Bernard, R.T.F., 2007. Habitat selection and feeding ecology of the cheetah (Acinonyx jubatus) in thicket vegetation: is the cheetah a savanna specialist?. Journal of Zoology, 271(3), pp.310-317.
[3] Shultz, S., Britnell, J.A. and Harvey, N., 2021. Untapped potential of physiology, behaviour and immune markers to predict range dynamics and marginality. Ecology and Evolution, 11(23), pp.16446-16461.
[4] Harvey Sky, N., Britnell, J., Antwis, R., Kartzinel, T., Rubenstein, D., Toye, P., Karani, B., Njeru, R., Hinchcliffe, D., Gaymer, J., Mutisya, S. and Shultz, S., 2024. Linking diet switching to reproductive performance across populations of two critically endangered mammalian herbivores. Communications Biology, 7(1), p.333.
[5] Harvey Sky, N., Jackson, J., Chege, G., Gaymer, J., Kimiti, D., Mutisya, S., Nakito, S. and Shultz, S., 2022. Female reproductive skew exacerbates the extinction risk from poaching in the eastern black rhino. Proceedings of the Royal Society B, 289(1972), p.20220075.
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Artificial Intelligence Methodology in Structural Biology
Publishing Model: Hybrid
Deadline: Nov 30, 2026
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in