Variation in the mu-opioid receptor gene (OPRM1) moderates the influence of maternal sensitivity on child attachment
Published in Behavioural Sciences & Psychology

Whether you happen to be a human or a rhesus macaque, life in infancy is precarious business.
Developmentally immature and unequipped to deal with a world teeming with dangers, infants rely entirely on a dedicated caregiver—like mom—to provide them with the nurturance and protection they need to survive. For the infant or young child, therefore, staying close to mom is, quite literally, a matter of life or death. The central idea of attachment theory (Bowlby, 1982) is that this vulnerability led to the evolution of a psychobehavioural system, called the attachment system, that regulates caregiver-seeking behaviour. The attachment system can be likened to a regulator—for example, a thermostat that serves to keep temperature constant. When the thermostat’s sensors detect deviation from the set-point, control mechanisms (heating, cooling) activate, bringing the temperature closer to the set-point, at which point the control mechanisms shut off.
In the case of the attachment system, the condition to be maintained is proximity to the caregiver, and the control mechanisms are proximity-seeking and -maintenance behaviours such as seeking, crying, and clinging. This proximity is best conceptualised in emotional terms, rather than units of physical distance. A child who feels that mom is engaged and emotionally present will experience a state of safety, comfort, and contentment that attachment researchers call felt security. Importantly, this state of felt security is critical to the pursuit of other activities essential for the child’s development, namely play and exploration of the physical and social world. In this sense, the caregiver (attachment figure) is said to serve as a secure base for the child, encouraging exploration by their mere availability.
We can see the attachment system clearly at work during a famous experimental paradigm, called the Strange Situation, developed by Mary Ainsworth (Ainsworth, Blehar, Waters, & Wall, 1978). In this paradigm, infants or young children visit the laboratory with their mother and go through a set of brief separations and reunions while researchers record and observe their behaviour. When the mother leaves the child alone in this unfamiliar environment, the child will typically exhibit distress. Losing all interest in the toys strewn about the room, the child will cry and search for mom (attachment system activation); when mom returns and consoles the child, however, the child will calm down and resume play. Importantly, some children deviate from this prototypical pattern. Of particular relevance to the present investigation, some children, described as ambivalent or anxious, not only exhibit high levels of distress when mom leaves, but also remain distressed upon her return. Unlike the prototypical secure children described above, ambivalent children are not soothed by contact with mom; they continue to cry and cling to her and are unable to resume play.
What accounts for this deviation from the typical pattern? Attachment theory argues that the child’s inability to derive comfort from and use the caregiver as a secure base stems from a history of interactions wherein the caregiver was less sensitive, failing to reliably and warmly attend to the child’s needs. It is thought that children characterised by ambivalence cope with maternal insensitivity by hyperactivating their attachment system, which manifests as heightened separation distress that persists beyond reunion and hypervigilance for any cues of attachment figure unavailability. For these children, it is as if the switch for the proximity-regulating attachment system is chronically stuck on “on”. Of note, there is a certain adaptive value to this pattern of behaviour—after all, if one’s caregiver is unreliable, it makes sense to ramp up one’s bids for attention to ensure that one’s needs will be met. However, in the long-term, attachment ambivalence may contribute to a host of difficulties with emotion regulation, interpersonal relationships, and even physical health (Mikulincer & Shaver, 2016), making it imperative to gain a better understanding of the risk factors for this pattern.

While maternal sensitivity has been reliably linked to attachment insecurity in children (De Wolff & van Ijzendoorn, 1997), it only explains part of the variance in children’s attachment behaviour. Our team wondered how genetic differences may interact with the caregiving environment to shape attachment. We were particularly intrigued by a study in rhesus macaques (Barr et al., 2008) focusing on a gene coding for the μ-opioid receptor (OPRM1 C77G). The μ-opioid receptor binds opioid drugs like morphine and mediates their rewarding and pain-relieving effects (Matthes et al., 1996), and it is thought that the rewards of social contact may rely on the same receptors involved in mediating the rewarding effects of morphine and other drugs (Panksepp, Siviy, & Normansell, 1985). Consistent with this, Barr et al. found that OPRM1 C77G moderates the effects of maternal separation on the infant’s attachment behaviour. After being subjected repeatedly to prolonged bouts of separation from their mothers, infant macaques carrying the minor G allele behaved somewhat like ambivalently-attached children, exhibiting progressively higher levels of separation distress and more persistent clinging to mom upon reunion, at the expense of socialising with their peers.

Inspired by Barr et al.’s (2008) study, we examined whether an analogous gene in humans, OPRM1 A118G, would similarly interact with the caregiving environment to predict attachment-related affect and behaviour in young children. Following Barr et al., we predicted that children carrying the minor G allele may be particularly predisposed to hyperactivation of the attachment system in response to less sensitive maternal care. To test this hypothesis, we drew on data from the Maternal Adversity, Vulnerability and Neurodevelopment (MAVAN) project, a community-based study of Canadian mothers and their children assessed across the child’s development (O’Donnell et al., 2014). Our sample consisted of 191 mothers participating with their children (n=223). Maternal sensitivity (Ainsworth, 1969) was assessed in an at-home visit when the child was 18 months old, during which the mother interacted with her child . Child attachment was assessed when the child was 36 months old using the previously described Strange Situation paradigm.
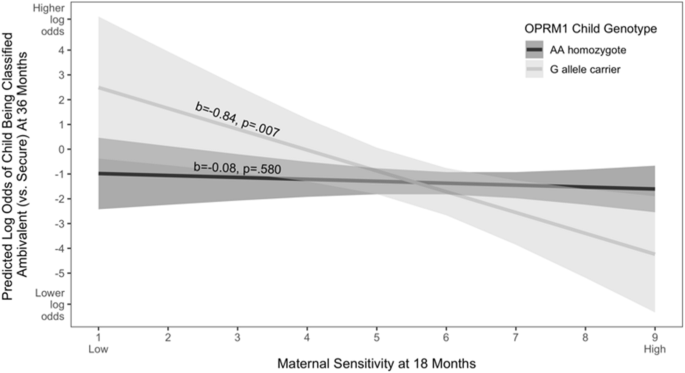
As one would expect, and consistent with prior research, maternal sensitivity was associated with attachment classification, with children of less sensitive mothers being more likely to be categorised as ambivalent (vs. secure). However, as hypothesised, child OPRM1 A118G genotype moderated this effect. Specifically, maternal sensitivity was associated with the likelihood of being categorised as ambivalent (vs. secure) for G allele carriers, but not for the AA homozygote group. That is, G allele carrying children were more likely to display ambivalence when their mother was previously rated as less sensitive (they were also less likely to display ambivalence when their mother was previously rated as more sensitive).
Results of multinomial logistic model predicting classification as ambivalent vs. secure as a function of OPRM1 child genotype and maternal sensitivity

Note. The X-axis reflects levels of maternal sensitivity measured when the child was 18-months old, with higher numbers indicating greater maternal sensitivity. The Y-axis reflects the predicted log odds of the child being classified as ambivalent (vs. secure) at 36-months, with higher numbers indicating greater log odds of being classified as ambivalent (vs. secure). Shaded areas represent 95% confidence intervals.
These results parallel those reported by Barr et al. in rhesus macaques. They are also consistent with earlier work by some of our team members conducted in adult romantic couples. In that study (Tchalova, Sadikaj, Moskowitz, Zuroff, & Bartz, 2021), romantic partners reported on their interactions with each other over a three-week period, specifically indicating how they had behaved in the interaction, how their partner had behaved, and how secure they felt towards their partner. Not surprisingly, participants felt less secure following interactions in which their partner reported behaving in a cold or quarrelsome manner (e.g., criticising, ignoring the other). Critically, however, OPRM1 A118G genotype again moderated this effect, with G allele carriers’ feelings of security being more strongly affected by their partner’s cold/quarrelsome behaviour.
Taken together, these studies suggest that the OPRM1 118G variant may predispose individuals to greater interpersonal sensitivity, and that they will be more strongly affected by the quality of their social interactions. More broadly, these findings are consistent with the theory that endogenous opioids play a role in regulating attachment-related affect and behaviour.
One important question concerns the implications of this gene for more serious and prolonged social separation, such as bereavement. As Barr and colleagues point out, there may be evolutionary value in the persistence exhibited by G allele carriers in eking out support from reluctant or distracted caregivers, particularly during times of environmental stress. However, such prolonged hyperactivation of the attachment system may have detrimental psychological and physiological effects when support is lost forever.
Finally, while our study had several methodological strengths (objective behavioural measures of both maternal sensitivity and child attachment, longitudinal assessment), the sample size was not large and the findings should thus be considered preliminary prior to replication. Further, replication in more diverse samples would help establish generalisability of the finding. Future research would also benefit from simultaneous examination of multiple neurochemical systems implicated in attachment (e.g., oxytocin, dopamine). Nonetheless, the current findings shed light both on the particular risk factors for attachment anxiety and, more broadly, the neurobiological underpinnings of attachment in humans.
References
Ainsworth, M. D. S. (1969). Maternal Sensitivity Scales; The Baltimore Longitudinal Project (1969). Baltimore, MD: Johns Hopkins University.
Ainsworth, M. D. S., Blehar, M. C., Waters, E., & Wall, S. N. (1978). Patterns of attachment: A psychological study of the Strange Situation. New York: Psychology Press. http://doi.org/10.4324/9780203758045
Barr, C. S., Schwandt, M. L., Lindell, S. G., Higley, J. D., Maestripieri, D., Goldman, D., … Heilig, M. (2008). Variation at the mu-opioid receptor gene (OPRM1) influences attachment behavior in infant primates. Proceedings of the National Academy of Sciences of the United States of America, 105(13), 5277–5281. http://doi.org/10.1073/pnas.0710225105
Bowlby, J. (1982). Attachment and loss. Vol I: Attachment (2nd ed.). New York, NY: Basic Books.
De Wolff, M. S., & van Ijzendoorn, M. H. (1997). Sensitivity and attachment: A meta-analysis on parental antecedents of infant attachment. Child Development, 68(4), 571–591. http://doi.org/10.1111/j.1467-8624.1997.tb04218.x
Matthes, H. W., Maldonado, R., Simonin, F., Valverde, O., Slowe, S., Kitchen, I., … Kieffer, B. L. (1996). Loss of morphine-induced analgesia, reward effect and withdrawal symptoms in mice lacking the mu-opioid-receptor gene. Nature, 383(6603), 819–23. http://doi.org/10.1038/383819a0
Mikulincer, M., & Shaver, P. R. (2007). Attachment in adulthood: Structure, dynamics, and change. New York, NY: The Guilford Press.
O’Donnell, K. A., Gaudreau, H., Colalillo, S., Steiner, M., Atkinson, L., Moss, E., … Meaney, M. J. (2014). The Maternal Adversity, Vulnerability and Neurodevelopment project: Theory and methodology. The Canadian Journal of Psychiatry, 59(9), 497–508. http://doi.org/10.1177/070674371405900906
Panksepp, J., Siviy, S., & Normansell, L. (1985). Brain opioids and social emotions. In Psychobiology of attachment and separation (pp. 3–49). New York: Academic Press.
Tchalova, K., Sadikaj, G., Moskowitz, D. S., Zuroff, D. C., & Bartz, J. A. (2021). Variation in the μ-opioid receptor gene (OPRM1) and experiences of felt security in response to a romantic partner’s quarrelsome behavior. Molecular Psychiatry, 26(8), 3847–3857. http://doi.org/10.1038/s41380-019-0600-4
Follow the Topic
-
Translational Psychiatry
This journal focuses on papers that directly study psychiatric disorders and bring new discovery into clinical practice.

Related Collections
With Collections, you can get published faster and increase your visibility.
Moving towards mechanism, causality and novel therapeutic interventions in translational psychiatry: focus on the microbiome-gut-brain axis
Publishing Model: Open Access
Deadline: Nov 15, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in