Functional hints from elusive motifs
Published in Bioengineering & Biotechnology

Recent times have seen exciting scientific progress, in particular in structural biology. We can solve the structure of huge proteins at a resolution level that sometimes we have the impression that we almost touch them, we can make numerical simulation at different theoretical levels on the most challenging molecules and also, we have been starting to be able to design new proteins almost from first principles. Yet, linking structures to function often remains challenging.
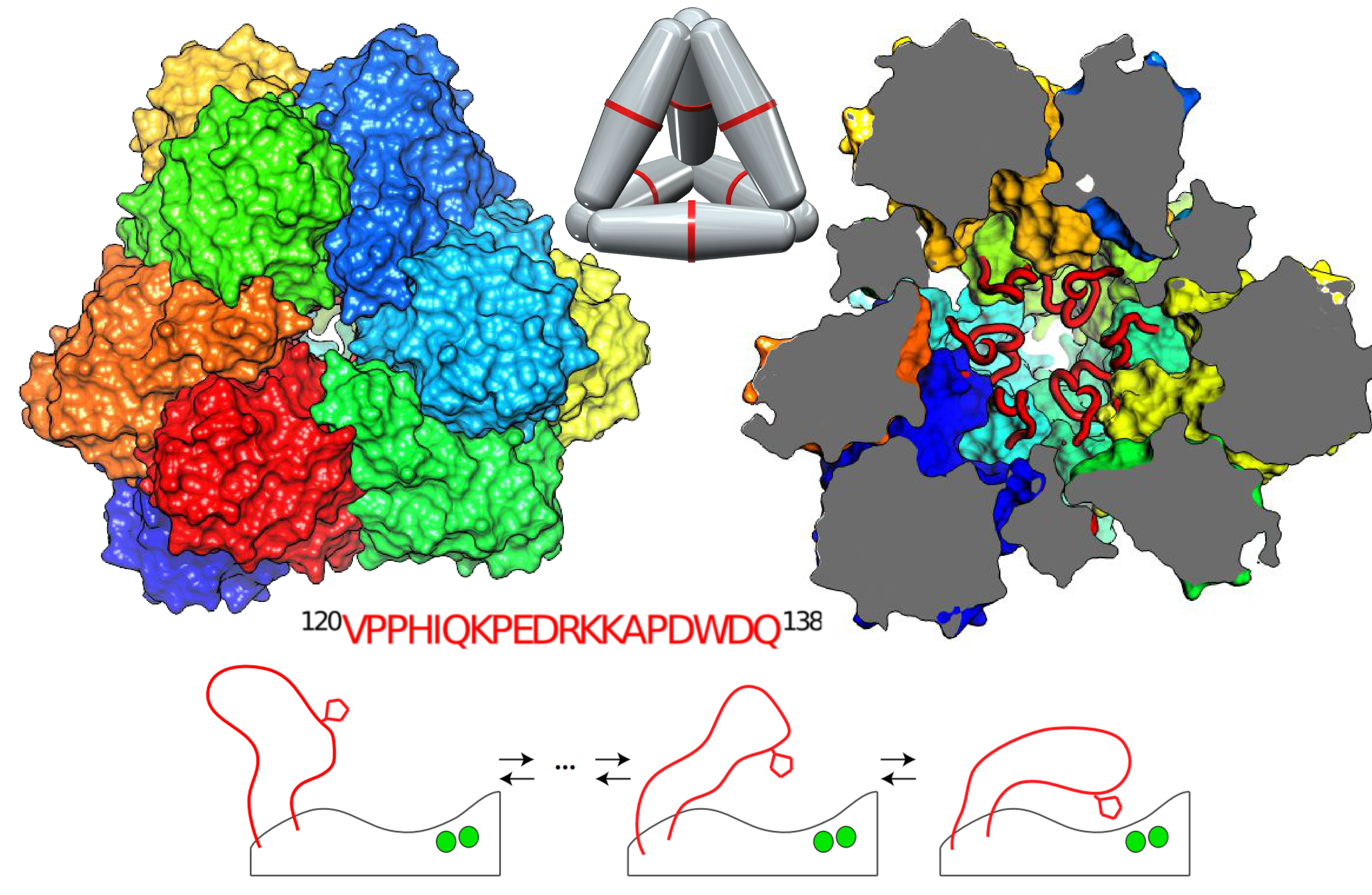
In our research, we aimed to decipher how a huge enzyme (TET2 aminopeptidase) is able to perform its proteolytic tasks and what is the key structural determinant that make it functional. Structural biologists generally deduce function from the 3D structures that e.g. crystallography provides. Hence, we jump to the PDB to look for its structures and luckily for us, there were a lot of nice structural details that emerge from the X-ray structure that allow us start to understand how this challenging protease might work. The TET2 aminopeptidase forms a dodecamer of around 0.5 MDa, with a well-defined entry pore for substrates selection and catalytic sites protected inside the chamber, that are the responsible to catalyse the peptide cleavage. TET2 is able to cleavage peptides that range from 4-40 aminoacids in length inside the wide catalytic chamber that is formed by the dodecameric assembly.
Looking in more details into the crystallographic protein structure reveals that the “wide open space” of the catalytic chamber is misleading – as long loop regions are missing in the crystallographic model. Our study started by realising that in functional tests with a protein that lack these “invisible” loops, the enzymatic activity is dramatically reduced. Hence, “invisible” (to crystallography) parts are essential to drive the enzyme activity.
We used magic-angle spinning NMR – a technique that can overcome problems that solution NMR generally has when studying proteins as large as TET2 – in order to decipher how important the missing structural sequence could be not only on TET2 but also different homologous that are part of the protein family. One more surprise are waiting for us: all the homologous present a high conserved residues “PPH” into the missing region and several residues inside the huge cavity are co-evolving.
So, there are something encoded in the protein function that the evolution trough the millions of try and error trials want to conserved. Of course, we were very encouraged now and we decide to touch the loop sequence on different key functional parts, and revealed which parts of this loop are central to function.
But we are structural biologist and we need the structure to solve the puzzle and now, CryoEM give us some hints around. At 100K a very weak electronic density allow us to model a loop structure close to the active site. Now we have a name for the elusive sequence: “The missing loop”.
And still, how can we probe the flexibility? and the answer will come from our well-loved NMR experiments. But we have just to confess, that the life it is never easy. After a lot of work of NMR sample preparation and switching between solution and solid state NMR experiments, the first results start to pop-up. And the NMR are really great for almost all the protein, except those residues that are part of the missing loop: Now, we have something in common with our X-ray crystallographers colleagues, nobody are able to make the missing loop visible!!
Sad story but not so sad because we have still a magic bullet on our hands (the first one). Although we are not able to see the loop, we can put in evidence from our NMR experiments all the environment that are in contact with it. In short, we compare NMR experiments of the protein WT and our mutant without the loop (and without enzymatic activity) and here we arrive to our second surprise: we can probe in an indirect way that the invisible loop are touching functional parts of the protein chamber that are very far in the space, revealing in this way, a high degree of mobility.
But as soon that we progress on this story, more questions are seeding on our heads that claim for answers. If the loop are moving, in which time scale are moving and how this motion are associated with the protein function? A very general answer could emerge from the NMR: as a rule of thumb, if you do not see the peaks, presumably is because the dynamic time scale of motion is in the microsecond regime and we lose all the spin magnetization by spin relaxation.
And now, our second magic bullet: We change the isotopically labelling scheme from fully labelled protein to only the methyl groups of the aliphatic aminoacids, then the magnetic relaxation should be less efficient.
And again, life was not easy but we got at least a good news: a peak corresponding to the methyl group of the Val120 (a residue at the begging of the invisible loop) crop out very clear in the NMR spectra. Hence, we can start to see at least, one of the leg of our invisible friend.
And now, if we can see how the leg are moving, we can understand how the rest of the body might follow this motion and most important, how the protein dynamics are linked with the protein function in this challenging and fascinating macromolecular assembly.
If you want to know how this story continue, I invited you to have a look for the rest of the details on our recent publication in Nature Communication [Gauto et al. Nat. Comm., 2022]
Interested in structural biology of protein cages and their encapsulated biomolecules. Ours swords are Solid state NMR and Molecular Modelling
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in