Location, Location, Location: How infection site dictates outcome during S. aureus biofilm infection
Published in Microbiology, Biomedical Research, and General & Internal Medicine

Perspective
Infection is an omnipresent threat. Evolutionary biologists often speak of biological arms races between organisms competing for a single niche, where species evolve capabilities to enable them not just to outcompete but to uniquely exploit the biological properties of a rival (or prey) organism. While this phenomenon conjures images of cheetahs and gazelles, each evolving to become progressively faster over countless generations in an effort to outrun each other, this same process also likely occurs in a different, more microscopic, niche- the tissues of the human body. Only, you are the gazelle; bacteria are the cheetah.
The Threat: Staphylococcus aureus
One example of a formidable (cheetah-like) bacterium is Staphylococcus aureus (S. aureus), which is capable of infecting virtually any tissue in the human body due, in part, to an arsenal of virulence factors that target the mammalian immune system (1). Often an asymptomatic colonizer of human skin (>30% of the population) (2), S. aureus can cause invasive disease in many sites in the body, such as deeper layers of the skin where it forms abscesses, the bloodstream resulting in sepsis, and the heart where it can establish endocarditis or colonize heart valves. Each of these infections is associated with varying degrees of morbidity and mortality that is aided by the arsenal of immune evasive virulence factors mentioned earlier. Luckily, at present, most S. aureus infections can be successfully treated with antibiotics. However, S. aureus is notorious for rapidly acquiring antibiotic resistance (3) and is currently threateningly close to acquiring widespread resistance to our so-called ‘antimicrobials of last resort’ (4, 5).
The Biofilm Advantage
One additional feature of S. aureus that presents a particularly difficult challenge for treatment is the ability to form a biofilm. Biofilm formation is a strategy that likely initially evolved to allow bacteria to survive in harsh, nutrient-limiting environments like hydrothermal vents, rocks, trees, or underwater surfaces (6, 7). Biofilms are communities of bacteria held together by a dense matrix of extracellular DNA, proteins, and polysaccharides that shield bacteria from their surroundings (6). Biofilm infections are commonly associated with surgical implants or native tissues and are problematic because of their ability to circumvent clearance by the immune system and antibiotics. The latter is explained by multiple factors including metabolic dormancy of a subpopulation of bacteria within the biofilm and impaired access of antibiotics to organisms buried deep within the biofilm (8). Currently, these infections can only be treated with additional surgeries to physically disrupt and/or remove the biofilm nidus (often along with the infected implant or tissue surface) combined with long-term antibiotic therapy to clear residual bacteria. Even with such aggressive treatments, infection recurrence is common and devastating (9-12).
The Motivation for this Study
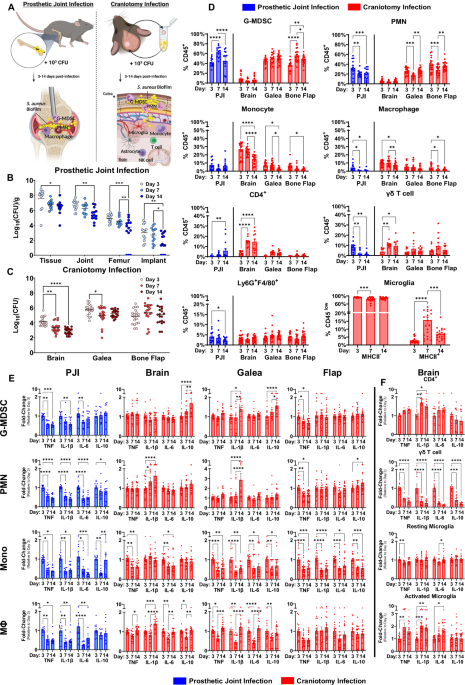
Our laboratory is working on new approaches to treat biofilms, particularly at uncovering ways to reprogram the immune system to better combat these infections and reduce the need for additional surgeries. However, before this can be done, we first need to understand the breadth of the problem. Biofilm can form following many different surgeries, including craniotomy and arthroplasty/prosthetic joint replacement (13-15). Our laboratory has extensively characterized the bacterial and immune responses during these infections in both animal models and patients revealing an anti-inflammatory bias. However, despite predictions that the local tissue microenvironment influences infection characteristics (16), no studies to date have directly compared immune responses at two different sites of biofilm infection. Our study addresses this gap in knowledge by comparing the functional, metabolic, and transcriptional characteristics of the immune response in two broadly similar models of biofilm infection, namely craniotomy and prosthetic joint infection (PJI). This is important, as future therapies would ideally be designed to target immune responses that are shared across a wide range of biofilm infections to maximize therapeutic efficacy.
Functional Differences During Alternative Biofilm Infections
When assessing PJI tissue and the galea (subcutaneous tissue above the skull from craniotomy infection), both showed largely similar bacterial burdens and leukocyte infiltrates at each timepoint assessed. These infiltrates were dominated by granulocytes, composed mainly of neutrophils (PMNs) and granulocytic myeloid-derived suppressor cells (G-MDSCs), a pathologically activated granulocyte subset with anti-inflammatory properties (17, 18). Despite these similarities, functional analysis revealed altered granulocyte activation during craniotomy or PJI. For example, in PJI, cytokine production was reduced in both PMNs and G-MDSCs as infection progressed, pathognomonic of chronic infection. However, during craniotomy infection PMNs and G-MDSCs revealed progressive elevations in IL-1β and IL-10 production, respectively. Therefore, granulocyte activation state is influenced by the biofilm microenvironment in a temporal fashion.
Metabolic Responses are Influenced by Biofilm Niche
Having identified differences in granulocyte activation by infection site, we hypothesized that the distinct tissue microenvironments in the joint or near the skull may contribute to differential granulocyte programming. It is well-known that metabolism can shape leukocyte polarization during infection (19, 20). Therefore, we next performed untargeted metabolomics on tissue extracts as well as granulocyte infiltrates from each infection site at multiple intervals post-infection. This approach revealed profound differences in the tissue metabolomes of craniotomy and PJI at every time point analyzed, along with corresponding alterations in intracellular metabolomes of granulocytes between infection modalities. Additionally, these metabolomes evolved over time, where large changes were observed longitudinally within a single infection site. Therefore, biofilm infections are associated with both temporal and spatial metabolic diversity which may underly the differences in granulocyte programming during craniotomy and PJI.
Enrichment analysis revealed that most differences across time or biofilm location were consistently observed in amino acid, fatty acid, and nucleotide metabolism. This provides broad direction for future experiments, as it is likely that metabolites in these categories have immunomodulatory effects, particularly with respect to granulocyte function during biofilm infection.
Transcriptional Differences are Shaped by Biofilm Niche
Having identified both functional and metabolomic heterogeneity between biofilm infections, we next assessed whether this extended to diverse transcriptional profiles in granulocytes isolated from craniotomy or PJI. Bulk RNA-seq identified significant alterations in metabolic genes and pathways such as fatty acid and organic acid metabolism, aligning with our metabolomic data. These findings were corroborated by scRNA-seq of leukocytes infiltrating craniotomy and PJI tissues of human subjects, which revealed immune pathways that were biased toward each infectious modality. This included PPAR signaling and PD-1/PD-L1 signaling in PJI, and IL-10 signaling and the integrated stress response in craniotomy infection, which were also detected in granulocytes from our mouse models.
Apples and Oranges
“This work establishes an important tenet regarding S. aureus biofilm infections, that is, no two tissue niches are identical…As future immunomodulatory therapies are developed to treat infection in the face of growing antimicrobial resistance, considerations related to the nature of the tissue niche and how this impacts leukocyte programming will need to be considered. A one-size-fits-all approach to immune modulation will not likely be effective as no microenvironment, and thus, no two biofilm infections are exactly equivalent.”
Put alternatively, biofilm infections are like apples and oranges (Figure 1).

Works Cited:
1. G. Y. C. Cheung, J. S. Bae, M. Otto, Pathogenicity and virulence of Staphylococcus aureus. Virulence 12, 547-569 (2021).
2. A. van Belkum et al., Co-evolutionary aspects of human colonisation and infection by Staphylococcus aureus. Infect Genet Evol 9, 32-47 (2009).
3. T. J. Foster, Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiology Reviews 41, 430-449 (2017).
4. A. Shariati et al., Global prevalence and distribution of vancomycin resistant, vancomycin intermediate and heterogeneously vancomycin intermediate Staphylococcus aureus clinical isolates: a systematic review and meta-analysis. Sci Rep 10, 12689 (2020).
5. M. Naghavi et al., Global burden of bacterial antimicrobial resistance 1990–2021: a systematic analysis with forecasts to 2050. The Lancet 404, 1199-1226 (2024).
6. K. Sauer et al., The biofilm life cycle: expanding the conceptual model of biofilm formation. Nature Reviews Microbiology 20, 608-620 (2022).
7. C. D. Rummel, A. Jahnke, E. Gorokhova, D. Kühnel, M. Schmitt-Jansen, Impacts of Biofilm Formation on the Fate and Potential Effects of Microplastic in the Aquatic Environment. Environmental Science & Technology Letters 4, 258-267 (2017).
8. T. K. Wood, S. J. Knabel, B. W. Kwan, Bacterial persister cell formation and dormancy. Appl Environ Microbiol 79, 7116-7121 (2013).
9. C. Gold, I. Kournoutas, S. C. Seaman, J. Greenlee, Bone flap management strategies for postcraniotomy surgical site infection. Surg Neurol Int 12, 341 (2021).
10. B. F. Ricciardi et al., Staphylococcus aureus evasion of host immunity in the setting of prosthetic joint infection: biofilm and beyond. Current reviews in musculoskeletal medicine 11, 389-400 (2018).
11. N. Sadhwani et al., Comparison of infection rates following immediate and delayed cranioplasty for postcraniotomy surgical site infections: results of a meta-analysis. World Neurosurgery 173, 167-175. e162 (2023).
12. C. E. Kandel et al., Predictors of Treatment Failure for Hip and Knee Prosthetic Joint Infections in the Setting of 1- and 2-Stage Exchange Arthroplasty: A Multicenter Retrospective Cohort. Open Forum Infectious Diseases 6, (2019).
13. Z. Van Roy et al., Single-cell profiling reveals a conserved role for hypoxia-inducible factor signaling during human craniotomy infection. Cell Reports Medicine.
14. J. Cheatle, A. Aldrich, W. E. Thorell, M. D. Boska, T. Kielian, Compartmentalization of immune responses during Staphylococcus aureus cranial bone flap infection. Am J Pathol 183, 450-458 (2013).
15. W. Zimmerli, A. Trampuz, P. E. Ochsner, Prosthetic-Joint Infections. New England Journal of Medicine 351, 1645-1654 (2004).
16. T. Bjarnsholt et al., The importance of understanding the infectious microenvironment. The Lancet Infectious Diseases 22, e88-e92 (2022).
17. S. Hegde, A. M. Leader, M. Merad, MDSC: Markers, development, states, and unaddressed complexity. Immunity 54, 875-884 (2021).
18. E. Medina, D. Hartl, Myeloid-Derived Suppressor Cells in Infection: A General Overview. J Innate Immun 10, 407-413 (2018).
19. C. M. Horn, T. Kielian, Crosstalk Between Staphylococcus aureus and Innate Immunity: Focus on Immunometabolism. Front Immunol 11, 621750 (2020).
20. M. A. Lynch, Can the emerging field of immunometabolism provide insights into neuroinflammation? Prog Neurobiol 184, 101719 (2020).
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Biosensing
Publishing Model: Hybrid
Deadline: Sep 30, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in